Los linfocitos B o células B son un tipo de linfocito y un componente esencial de la respuesta inmune humoral. Los linfocitos B son un componente esencial del sistema inmunitario adaptativo. Las funciones principales de los linfocitos B son:
- Fabricar anticuerpos contra antígenos
- Funcionar como células presentadoras de antígeno (APC).
- Eventualmente transformarse en células de memoria B después de ser activados por la interacción con un antígeno.
Tienen dos procesos de maduración y diferenciación:
1.- El primero se produce en la médula ósea, que no está expuesta a los antígenos.
2.- El segundo proceso de maduración se produce en el ganglio linfático. En esta segunda etapa la célula B se diferencia en otros tipos de células: células plasmáticas y células B de memoria.
DESARROLLO Y DIFERENCIACIÓN
Maduración independiente del antígeno
Es la fase que se produce en la médula ósea. Aquí se producen todas las células sanguíneas del cuerpo.
Maduración dependiente del antígeno
Se produce en el ganglio linfático. Se produce después de la activación de células B mediante la unión de antígenos y co-estimulación. Estas señales promueven la diferenciación de los linfocitos B en células plasmáticas y células B de memoria.
Las células plasmáticas secretan anticuerpos que son responsables de la neutralización de antígenos mediante la unión a ellos, de modo que se conviertan en objetivos más fáciles para los fagocitos. Las células plasmáticas mueren a los pocos días. Los anticuerpos permanecen en el sistema durante un poco más de tiempo.
Las células B de memoria se forman específicamente contra antígenos encontrados durante la respuesta inmune primaria; ya que pueden vivir mucho tiempo. Las células de memoria permanecen en el cuerpo durante mucho más tiempo que las células plasmáticas y anticuerpos, a menudo años. Son importantes para proporcionar inmunidad, ya que pueden responder rápidamente durante una segunda exposición a su antígeno específico.
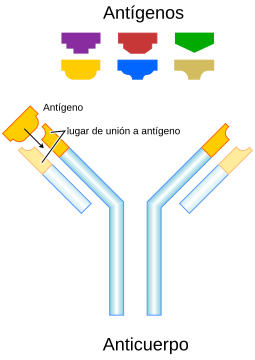
Cuando se activa un linfocito B, produce anticuerpos, unas proteínas que luchan contra antígenos. Los anticuerpos son específicos para un único antígeno, por lo que hay muchos tipos de células B en el cuerpo.
Todas las células plasmáticas que descienden de una sola célula B producen el mismo anticuerpo que está dirigido contra el antígeno que estimula la maduración. El mismo principio se aplica con las células B de memoria. Por lo tanto, las células plasmáticas y células de memoria “recuerdan” el estímulo que dio lugar a su formación
Ciclo de vida de los linfocitos B

Poblaciones de los linfocitos B
· Linfocitos B2: 95% de los linfocitos B en sangre y ganglios linfáticos
· Linfocitos B1: predominan en las cavidades peritoneal y pleural
· Linfocitos B de la zona marginal del bazo
BIBLIOGRAFÍA
E. Fernández Mastache, A. González Fernández, S. Lorenzo Abalde, Enfermedades del sistema inmune, Universidad de la Rioja/ España, 2001-2016
Mauri, Claudia; Bosma, Anneleen, “Immune Regulatory Function of B Cells”. Annual Review of Immunology,University College London, Janualy 3 2012.
http://www.fmed.uba.ar/depto/microbiologia/TE%C3%93RICO%20N%C2%B05.pdf