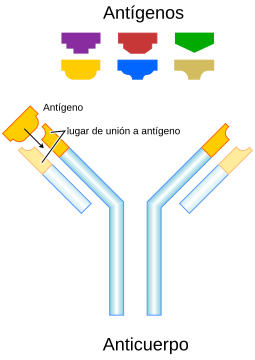
En la respuesta inmune participan células y moléculas
distribuidas por todo el organismo, tanto en la circulación
como en los diversos tejidos del organismo. Sin embargo,
grupos de células inmunocompetentes conforman tejidos
especializados del sistema inmune, los que a su vez se integran
como órganos linfoides.
Los órganos linfoides se dividen funcionalmente en dos
tipos:
• Primarios o centrales: Son aquéllos en los que los linfocitos
se originan y maduran, a través del mecanismo de
linfopoyesis y/o la adquisición de las características
que los capacitan a responder ante un antígeno extraño.
En este sitio las células que actúan contra estructuras
moleculares propias son eliminadas y sobreviven.
Médula ósea

En este órgano se generan las células troncales hematopoyéticas
(stem cells) o células madre, origen de todas las
células sanguíneas. En la vida fetal emergen inicialmente
del saco embrionario y posteriormente del hígado y del bazo;
al nacimiento, la médula ósea se convierte en el principal
centro hematopoyético. Si la demanda de células es muy
grande o hay daño medular, se perfilan como auxiliares en
la hematopoyesis, el hígado y el bazo.
La médula ósea se encuentra en el interior del hueso como
una estructura reticular inmersa entre grandes trabéculas, en
cuyos espacios se encuentran adipocitos, fibroblastos del
estroma y precursores de las células sanguíneas. En los procesos
de crecimiento y diferenciación de las células progenitoras,
participan una variedad de factores estimuladores,
entre los que se encuentran las citocinas: IL-1, 3 (acción
multilineal), 6,7 (línea linfoide), 11 (generación de plaquetas)
y factores estimuladores de colonias de granulocitos y
monocitos (GM-CSF, G-CSF).
Durante la diferenciación del linfocito B participan activamente
las células del estroma con la liberación de citocinas
y factores de crecimiento; en esta etapa, las células B
que muestran autorreactividad son disminuidas por apoptosis,
lo que sucede aproximadamente en 50%. Finalmente, el
linfocito B maduro emerge de la médula y a través de la
circulación, se dirige a los órganos linfoides secundarios
para ejercer su función efectora.
El linfocito T que también se origina en la médula, sale
de ella inmaduro (timocito). A continuación, el timocito
guiado por señales quimioatractantes generadas por quimiocinas
en el timo, ingresa a este órgano para completar su
desarrollo y adquirir las características de madurez que lo
facultan para responder a un antígeno.
Timo
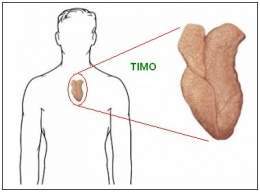
Es un órgano bilobulado, situado en la parte anterior del
tórax. Cada lóbulo se divide por trabéculas de tejido conjuntivo
en pequeños lóbulos constituidos por varias zonas.
En la corteza se encuentran células epiteliales también llamadas
nodriza (nurse) que interaccionan con los timocitos
proporcionándoles, al igual que los otros tres tipos de células
epiteliales, hormonas tímicas (timosina, timopoyetina,
factor tímico sérico) que les ayudan a madurar. Más profundamente,
las células epiteliales forman una densa malla, que
el timocito cruza para finalmente llegar a la médula en donde
se encuentran corpúsculos de Hassall (los que recientemente
han sido implicados en la formación de la célula CD4CD25),
macrófagos, escasas células mioides secretoras de citocinas
(IL:1, 3, 6, 7) y células dendríticas interdigitantes (ricas en
MHC) con las que establece contacto e interactúa.

Se produce así, la selección positiva, lo que significa que
después de haber sido sometidos a un escrutinio por diferentes
células, sólo sobreviven aquellos timocitos que no son
autorreactivos; se propicia el desarrollo de los linfocitos
capacitados para reaccionar frente a moléculas extrañas y se
induce la apoptosis de los que muestran afinidad por molé-
culas propias.
Finalmente, los linfocitos maduros con sus marcadores
CD4 (Th- linfocito cooperador) o CD8 (Tc- linfocito citotóxico)
salen del timo a través de las vénulas con endotelio
columnar alto y entran al torrente sanguíneo en donde muchos
de ellos se quedan (el 75% de los linfocitos circulantes
son T), el resto se dirige a los órganos linfoides secundarios
para ejercer el reconocimiento específico del antígeno correspondiente.
El timo desarrolla su máxima actividad durante los primeros
años de vida, lo que se refleja en la producción de
linfocitos que, en un individuo de 35 años de edad, corresponde
al 20% de la generada en el neonato. Conforme avanza
la edad, disminuye el número de linfocitos T vírgenes,
por lo que la respuesta inmune celular en etapas avanzadas
depende, principalmente, de los linfocitos T de memoria.
• Secundarios o periféricos: Son estructuras especializadas
en la recolección de antígenos de distintos compartimentos
anatómicos. En ellos se lleva a cabo la activación
de los linfocitos maduros, a través de la «presentación»
o el contacto con el antígeno, lo que da inicio a la respuesta
inmune específica, con la consiguiente proliferación
clonal y la generación de células de memoria.
Ganglio linfático
El ganglio forma parte del sistema linfático que filtra por
zonas los antígenos procedentes del líquido intersticial y de
la linfa. Los antígenos libres o las células portadoras de los
antígenos pueden penetrar al ganglio por los ductos denominados
vasos linfáticos aferentes, para establecer contacto
con los linfocitos ubicados en él. Los linfocitos sanguí-
neos llegan al ganglio principalmente por vía hematógena a
través de vénulas.
El ganglio está rodeado por una cápsula de tejido conectivo
y estructurado por tres regiones. En la corteza predominan
las células B y se localizan los agregados celulares denominados
folículos primarios. En la paracorteza abundan
los linfocitos T y las células dendríticas interdigitantes
que dan soporte y poseen moléculas MHC II, por lo que
actúan principalmente como presentadoras. En la médula
del ganglio hay macrófagos, linfocitos T, B y numerosas
células plasmáticas.
En el folículo primario abundan las células B, hay algunos
linfocitos T y células dendríticas foliculares de soporte,
cuyas prolongaciones circunscriben a los linfocitos. Si el
antígeno penetra libremente puede ser captado directamente
por el linfocito B o por el macrófago; por otra parte, si el
antígeno es transportado por un fagocito, puede ser presentado
al linfocito T o B. En ambos casos existe la posibilidad
inmediata de que el linfocito B o T se active, lo que conlleva
a un aumento en su tamaño (principalmente si se activa B
y se transforma en célula plasmática) y a un incremento en la
actividad del retículo endoplásmico generador de proteínas
(anticuerpos, citocinas). Como consecuencia, debido a los
cambios que conlleva la activación de los linfocitos éste se
transforma en folículo secundario o centro germinativo de
Flemming.
Los centros germinales corresponden a zonas
con células en intensa proliferación, que originan células
efectoras y de memoria, localizándose en ellos linfocitos B,
macrófagos, células dendríticas, plasmáticas y algunos linfocitos
T.
Bazo
Es un órgano situado en el hipocondrio izquierdo con un
peso aproximado de 150 g. Tiene dos tipos de tejidos, el que
corresponde a la pulpa blanca está constituido por una arteriola
central cubierta con una vaina de tejido linfoide periarteriolar,
los linfocitos T se encuentran alrededor del vaso
sanguíneo y las células B confluyen y forman folículos primarios.
En el sitio de transición entre ambas zonas hay un
gran número de macrófagos que presentan antígenos a los
linfocitos y fagocitan células deterioradas, principalmente
eritrocitos.
La otra zona del bazo denominada pulpa roja
está integrada por sinusoides vasculares que finalmente conectan
con la vena esplénica, lo que permite la salida de la
sangre que ingresa, constantemente, a través de la arteria.
El bazo filtra sangre de manera similar a como los ganglios
filtran linfa, y este mecanismo es uno de los más efectivos
para depurar al organismo de gérmenes que de alguna
manera llegan a la circulación. Cada día la mitad del volumen
sanguíneo corporal total pasa por este órgano y en él se
lleva a cabo la fagocitosis, no sólo de antígenos sino también
de células senescentes o dañadas.
Tejido linfoide asociado a mucosas (MALT)
Son agrupaciones linfoides no capsuladas situadas en
áreas submucosas. Las células de cada sitio tienen
distintos fenotipos y características funcionales. La
mayoría de los linfocitos intraepiteliales son T con predominio
del tipo CD8; en humanos aproximadamente el 10%
corresponde a linfocitos intraepiteliales T caracterizados
por su capacidad para responder directamente ante cualquier
antígeno. Al igual que en el resto de los órganos descritos,
hay tejido linfoide organizado en folículos primarios con
abundantes linfocitos B y centros germinales o folículos secundarios.
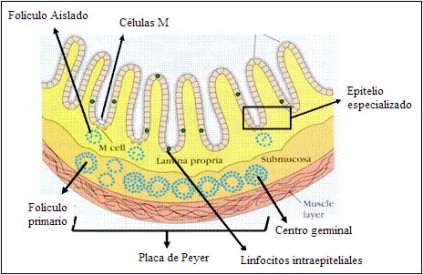
La inmunidad generada en estos sitios se enriquece
con la actividad desplegada por la gran cantidad de anticuerpos
(IgA), que se encuentran inmersos en las mucosas.
Entre los órganos linfoides secundarios, existen variaciones
tanto en la estructura como en la forma de llegada de
un antígeno, así al ganglio llega principalmente por la linfa,
en el bazo por la sangre y en algunos tejidos asociados a
mucosas a través de células. Sin embargo, el proceso desencadenado
por la interacción antígenos-células linfoides, en
los folículos primarios, descrito en la sección correspondiente
al ganglio, se realiza de manera similar en los diferentes
órganos secundarios.
Los linfocitos generados en la médula ósea y el timo recirculan
por todo el organismo, a través de la sangre y la
linfa, para finalmente ingresar a los órganos linfáticos secundarios
y establecer contacto con el antígeno correspondiente.
Muchos linfocitos ya activados o de memoria, salen
de los órganos linfoides y se ubican en el sitio de entrada del
antígeno con el que interaccionaron o migran a sitios de
inflamación e infección.
Otros, principalmente los de tipo T
permanecen en la circulación, adheridos a las paredes de los
vasos sanguíneos o dispersos en los tejidos linfoides asociados
a mucosas, en compañía de los B, que se localizan principalmente
en los órganos linfoides. Esta distribución estratégica
muestra la amplia cobertura del sistema linfoide, para
establecer una óptima vigilancia y protección específica a
través de todo el organismo.
BIBLIOGRAFÍA
1. Ferreira A, Afani S, Lanza B, Aguillón J, Sepúlveda C. Inmunología
básica y clínica. Ed. Mediterráneo, Santiago, Chile,
2005.
2.- Gloria Bertha Vega Robledo, Inmunología para el médico general
Órganos linfoides, Rev Fac Med UNAM Vol. 52 No. 5 Septiembre-Octubre, 2009